
| JASNH, 2002, Vol. 1, No. 3, 43–48 | Copyright 2002 by Reysen Group. 1539-8714 |

This report investigates if a partial loss of immunity results in alterations of murine behaviors in the open field (ambulation, a correlate of exploration or extroversion, and defecation, a correlate of neuroticism); positive findings would implicate the immune system in the expression of the above behaviors. Twelve female mice and 12 male mice, of the outbred CD1 strain, received a daily injection of the immunosuppressive drug methotrexate, at 3 mg/kg/injection, for several days, whereas 10 female mice and 9 male mice received an injection of saline. Once immunodepression was established, as revealed by impaired antibody responses to rat erythrocytes, the mice were tested in the open field, where ambulation, rearing, defecation, and grooming were recorded. The same behaviors were recorded before the course of methotrexate (baseline). Although methotrexate decreased the antibody response (i.e., it produced immunodepression), it did not affect the behaviors, which points to little participation of the specific immunity on the recorded behaviors. |
One issue in psychoneuroimmunology is the relation between immunity and behavior. There are two broad forms of immunity: natural (or innate) and acquired (or specific). Activation of innate immunity, following infection by pathogenic microorganisms, results in behavioral alterations (sickness behavior; Berczi & Szentivanyi, 1996; Dantzer, Bluthé, Kent, & Goodall, 1993; Maier & Watkins, 1998), but the effect of specific immunity on behavior is far less studied.
The effect of specific immunity on behavior may be investigated in two ways: either by generating the specific immune response (for instance, by inoculation of an innocuous vaccine) and checking its effect on behavior (Gates et al., 1992; Vidal, 1999; Zacharko et al., 1997), or by suppressing the immune response and seeing how this maneuver alters behavior. The former approach has yielded diverse results; thus, elicitation of specific immunity decreases fear in the sheep (Gates et al., 1992), decreases pleasure in the mouse (Zacharko et al., 1997), or does not alter some consistent murine behaviors in the open field (Vidal, 1999). The latter approach (effect of diminished immunity on behavior) has been scarcely used (to my knowledge). In one report (Barnard, Behnke, Gage, Brown, & Smithurst, 1997a), mice immunodepressed by injection of anti-thymocyte serum displayed less motor activity and aggression than normal mice. In another report, by the same authors (Barnard, Behnke, Gage, Brown, & Smithurst, 1997b), male mice exposed to female odors and immunodepressed by injection of anti-thymocyte serum displayed similar motor activity and aggression than control mice (in the former report, the males were shielded from female odors). Males were used in both reports, but it would be interesting to know if loss of immunity also affected behavior of females; consequently, this paper reports results obtained in female mice.
If loss of immunity affected behavior, then the immune system would be implicated in the expression of the affected behaviors, and if those behaviors were stable, then the immune system might be implicated in the expression of traits (ambulation of rats in the open field has been considered a correlate of extroversion, and defecation a correlate of neuroticism (Garcia-Sevilla, 1984), although ambulation in the open field could also be a correlate of exploration (Simmel & Bagwell, 1983)). The results reported here show that a moderate, and presumably transient, depression of the specific immunity did not alter some murine behaviors in the open field (ambulation, rearing, defecation, and grooming). This result suggests that the immune system contributes minimally to the expression of those behaviors.
Subjects. CD1 mice of both sexes were purchased from Charles River (CRIFFA, Barcelona, Spain). Those mice were mated in our laboratory, and their offspring were the subjects for the experiments reported here. The mice, housed 3-4 mice per cage, lived under a 12-hr light-dark cycle: the lights were on from 8:00 hr to 20:00 hr. The mice received food and water ad libitum. The temperature of the room was 22 ± 1 °C. At the beginning of the experiment, the mice were 2-3 months old.
Open field tests. Two open fields were used: the first one (square open field) was used to record basal behaviors (i.e., behaviors before immunodepression); the second one (circular open field) was used to record behaviors while the mice were immunodepressed.
Open field one was a (24 cm × 24 cm × 14 cm) square open field whose floor was marked off by diagonal lines in four triangles; its color was black. Open field two was a circular yellow surface, of 40 cm diameter, with a circular green wall 30 cm high; the floor was marked off by black lines in 16 sectors. During 4 min. and 30 s, the following behaviors were recorded from each mouse: ambulation (number of lines crossed by the mouse), rearing (number of times the mouse stood completely erect on its hind legs), defecation (number of fecal boli), and grooming (number of times the mouse scratched its face with its forepaws). The open field was wiped clean with soapy water before each mouse was placed in it; a different soap was used with each open field. Open fields 1 and 2 were placed in different rooms.
Immunodepression. The immunodepressant drug methotrexate (Sigma) was suspended in saline and enough 0.2 M sodium hydroxide was added, with stirring, to bring the pH to 7.5; on the days specified below (Tables 1-2), 0.10 ml of this solution was injected subcutaneously into each mouse. The mean dose was 3 mg/kg/injection.
Immunization and antibody measurement. Before the course of methotrexate (MTX), the mice were injected intraperitoneally (ip.) with goat albumin (2 mg/mouse in saline), bled 8 days later, and the plasma level of (IgM and IgG) antibodies to goat albumin was measured by DIG-ELISA (diffusion-in-gel enzyme-linked immunosorbent assay). DIG-ELISA was as originally described (Elwing, Lange, & Nygren, 1980) with these modifications: a) the petri dishes were 13 cm in diameter, b) each petri dish contained 20 ml of 1% agar with 1 ml of 2% gelatin, c) development of colored spots was achieved by addition of a solution of diaminobenzidine, nickel chloride, and hydrogen peroxide, followed by further intensification of the ensuing color with silver (Ludány et al., 1993). The level of antibody to a given antigen in a test serum is expressed as a fraction of the level in a standard serum (standard sera for IgM and IgG were different).
During the course of MTX, and to verify immunodepression, the mice were injected ip. with rat erythrocytes (4.8 × 107 erythrocytes per female mouse; 3.9 × 107 erythrocytes per male mouse) and, 9 days later for females, or 7 days later for males, the mice were boosted (ip.)with rat erythrocytes (4.2 x 107 erythrocytes per female; 4.4 × 107 erythrocytes per male). The red blood cells came from a Long-Evans rat. Plasma levels of (IgM and IgG) antibodies to rat erythrocytes were measured by DIG-ELISA on a monolayer of erythrocyte stromata.
The mice were bled from the retroorbital plexus while they were anesthetized with ether.
Design. The (male or female) mice of each litter were randomly assigned to the treatment group or to the control group. Prior to the administration of MTX (baseline), the mice were assessed on immunity (antibody response to goat albumin) and in open field number one; during treatment (injection of MTX), the mice were assessed first on immunity (antibody response to an injection of rat erythrocytes) and afterward in open field number two. Tables 1 and 2 show the sequence of operations for female and male mice. Two different antigens were injected (goat albumin to assess basal immunity, and rat erythrocytes to assess immunodepression by MTX) because MTX is less efficient on antigen-primed lymphocytes than on naive lymphocytes (Winkelstein, 1994).
Two different open fields were used to avoid carry over effects from one session to the next in the same open field.
Statistics. The design was a pretest-posttest design (i.e., a between-within design with a baseline); in this case, the effect of treatment (MTX vs. saline), as well as the “treatment × occasion” interaction, is assessed by analysis of covariance (ANCOVA), with the measurement at baseline as the covariate (Edwards, 1985). In the experiment with females, some variables (grooming on day 37 and all IgM and IgG antibody measurements) did not meet the homogeneity-of-variances requirement (Cochran C > 0.83; p < 0.015), which precluded ANCOVA. Therefore, the effect of MTX on grooming, on the IgM antibody level, and on the IgG antibody level was assessed by rank ANCOVA (Huitema, 1980), whereas the effect of MTX on the other variables was assessed by conventional ANCOVA. The assumption of homogeneity of slopes was always met. Because 6 analyses of covariance were performed (one for each variable), the significance level was adjusted by Simes's correction (Wilcox, 1987) so that the experimentwise type I error probability was 0.05.
In the experiment with males, all the variables complied with Cochran's test of homogeneity of variances and with the homogeneity of slopes test; therefore, the “treatment x occasion” interaction and the “treatment” effect were assessed by ANCOVA. Because 5 analyses of covariance were performed (one for each variable; levels of IgM antibody were not measured), the significance level was adjusted by Simes's correction so that the experimentwise type I error probability was 0.05.
Before ANCOVA, I checked, by the Mann-Whitney U test, that control and experimental groups were comparable in every variable recorded at baseline.
The statistical package used was Statistica 5.5, '99 edition (Satsoft, Tulsa, Oklahoma).
Equivalence at baseline of experimental and control groups. A Mann-Whitney U test on every variable recorded at baseline (IgM and IgG antibodies to goat albumin, ambulation, rearing, defecation, and grooming in the open field) did not reveal statistically significant differences between the control and the experimental group (0.13 < p < 0.89), either in males or in females; the same results are also apparent by visual inspection of Figures 1-3: the 95% confidence intervals of control and experimental groups overlap.
Immunodepression by methotrexate. Injection of MTX decreased the antibody response of female mice to rat erythrocytes (Figure 1), and this result was confirmed by rank ANCOVA: for IgM, the “treatment × occasion” interaction was not significant (F(1,20) = 2.93; p = 0.10), but the treatment effect was so (F(1,19) = 19.10; p = 0.0003); for IgG, the results were similar: non significant “treatment × occasion” interaction (F(1,20) = 0.0009; p = 0.97) and significant treatment effect (F(1,19) = 68.23; p < 0.000001). Therefore, methotrexate depressed the antibody response of female mice.
Methotrexate also decreased the IgG antibody response of male mice to rat erythrocytes (Figure 3): ANCOVA revealed a significant “treatment × occasion” interaction (F(2,36) = 10.78; p = 0.0002) and a contrast on days 22, 31, and 48 showed that MTX-treated mice had less antibody than saline-injected mice (on day 22, F(1,18) = 9.65, p = 0.006; on day 31, F(1,18) = 10.33, p = 0.005; on day 48, F(1,18) = 12.84, p = 0.002). The level of IgM antibodies was not measured.
Effect of methotrexate administration on behaviors in the open field. Administration of MTX to female mice did not alter significantly any of the behaviors: this result is seen in Figure 2, which shows overlapping confidence intervals of experimental and control groups for every behavior, and is confirmed statistically: ANCOVA for rearing yields non significant “treatment × occasion” interaction (F(1,20) = 1.77, p = 0.20) or treatment effect (F(1,19) = 0.06, p = 0.81); ANCOVA for ambulation yields the same results (“treatment × occasion” interaction: F(1,20) = 1.45, p = 0.24; treatment effect. F(1,19) = 0.016, p = 0.90); ANCOVA for defecation also reveals non significant “treatment × occasion” interaction (F(1,20) = 2.21; p = 0.15) or treatment effect (F(1,19) = 0.11; p = 0.74), and rank ANCOVA for grooming shows a non significant “treatment x occasion” interaction (F(1,20) = 0.075; p = 0.79), and a trend toward significance for the treatment effect (F(1,19) = 4.35; p = 0.051); nevertheless, the treatment effect for grooming should not be declared significant after adjustment of the significance level by Simes's correction (Wilcox, 1987).
Methotrexate did not alter significantly the same behaviors in male mice. This is shown in Figure 3 and the ANCOVA results: for ambulation, neither the “treatment × occasion” interaction (F(1,18) = 0.031; p = 0.86) nor the treatment effect (F(1,18) = 0.008; p = 0.93) was significant; for defecation, the “treatment × occasion” interaction (F(1,18) = 0.44; p = 0.51) and the treatment effect (F(1,18) = 2.54; p = 0.13) were not significant; for grooming, neither the “treatment × occasion” interaction (F(1,18) = 0.40; p = 0.53) nor the treatment effect (F(1,18) = 0.04; p = 0.84) was significant; for rearing, the “treatment × occasion” interaction was significant (F(1,18) = 6.85; p = 0.017), but a contrast on day 27 (F(1,18) = 1.76, p = 0.20) and on day 50 (F(1,18) = 0.37, p = 0.55) failed to reveal statistically significant differences between rearing of saline-injected mice and MTX-injected mice (besides, the treatment effect did not reach significance (F(1.18) = 0.13, p = 0.72)).
This report shows that methotrexate decreases the specific immunity (Figures 1, 3, and Results) while leaving some behaviors unaltered (Figures 2-3). This finding suggests that a partial loss of immunity does not affect those behaviors. Yet, an alternative explanation could be that methotrexate cancels the putative effect of immunodepression on behavior. I think this interpretation is unlikely because methotrexate did not display one of its main side effects (leukopenia) at the dose used, and for the time spent, in this report (results not shown).
When investigating the effect of diminished immunity on behavior, depression of immunity should be moderate and transient. There are two reasons for this requirement: a) profound and prolonged immunosuppression increases the chances of infection, which would result in behavioral alterations and would confound interpretation of results, b) administration of large doses of immunosuppressive drugs, needed to achieve deep immunosuppression, is accompanied by side effects that may confound interpretation of results (for instance, a high dose of cyclosporine A, 20 mg/kg, increases defecation and decreases ambulation in the open field (von Horsten et al., 1998): whether those behavioral alterations are due to loss of immunity or to side effects is not clear). Because of these considerations, I found, by pilot experiments, that methotrexate, at 3 mg/kg/day, for up to 16 days, did not cause apparent side effects (the mice neither lost weight nor decreased the number of blood leukocytes; results not shown); besides, clinical experience shows that immunity recovers upon discontinuation of methotrexate.
The negative results reported here seem at variance with the results of another report (Barnard et al., 1997a) that described the decrease of activity and aggression, in male mice, after immunodepression by anti-thymocyte serum. Some differences between the experiment of Barnard et al. and mine could explain the varying results: a) the mouse strains are different, b) in the experiment by Barnard et al., locomotor activity is a composite of several motor activities (ambulation, climbing, digging in sawdust, sniffing the cage), c) according to Barnard and collaborators, when sawdust from cages that housed females was introduced into the male cages, immunodepression by anti-thymocyte serum did not alter behavior (Barnard et al., 1997b); in my experiment, male and female mice were caged in the same room, and the scent from females may have reached the males.
When working with females, one has to consider the putative contribution of the estrous cycle to the results. In the present experiment, the question is: could the difference in the estrous cycle of the females in the experimental and control groups explain the negative results?. This seems improbable because a) the assignment of females to both groups was random, which maximizes the chances that females in all phases of the sexual cycle are in both groups, b) at baseline (i.e., before methotrexate administration), both groups were equivalent in every behavior (Results), c) in male mice of the same strain, immunodepression by methotrexate did not alter the same behaviors (Figure 3 and Results).
The results reported here suggest that a moderate decrease of the specific immunity does not affect, in the mouse, the traits of extraversion and neuroticism (if ambulation and defecation in the open field are considered correlates of those traits; Garcia-Sevilla, 1984). Yet a comment is needed: the sample size used here (about 10 subjects in the control group and 12 in the experimental one) may be insufficient for weak effects to reach statistical significance. Therefore, the inference from this report is that a moderate loss of the specific immunity may not affect, or may affect weakly, the expression of some murine behaviors.
This work was supported by a grant (DGICYT No. PB95-0458) from the Spanish Ministry of Education.
Barnard, C.J., Behnke, J.M., Gage, A.R., Brown, H., & Smithurst, P.R. (1997a). Modulation of behaviour and testosterone concentration in immunodepressed male laboratory mice (Mus musculus). Physiology and Behavior, 61, 907-917.
Barnard, C.J., Behnke, J.M., Gage, A.R., Brown, H., & Smithurst, P.R. (1997b). Immunity costs and behavioural modulation in male laboratory mice (Mus musculus) exposed to the odours of females. Physiology and Behavior, 62, 857-866.
Berczi, I., & Szentivanyi, A. (1996). The pituitary gland, psychoneuroimmunology, and infection. In: H. Friedman, T.W. Klein, & A.L. Friedman, (Eds.), Psychoneuroimmunology, stress, and infection (pp. 79-109). Boca Raton, Florida: CRC Press.
Dantzer, R., Bluthé, R.M., Kent, S., & Goodall, G. (1993). Behavioral effects of cytokines: An insight into mechanisms of sickness behavior. In: E.B.de Souza, (Ed.) Neurobiology of cytokines (pp. 131-150). San Diego, California: Academic Press.
Edwards, A.L. (1985). Multiple regression and the analysis of variance and covariance (2nd ed.). New York: Freeman.
Elwing, H., Lange, S., & Nygren, H. (1980). Diffusion-in-gel enzyme-linked immunosorbent assay (DIG-ELISA): Optimal conditions for quantitation of antibodies. Journal of Immunological Methods, 39, 247-256.
Garcia-Sevilla, L. (1984). Extraversion and neuroticism in rats. Personality and Individual Differences, 5, 511-532.
Gates, G.R., Fell, L.R., Lynch, J.J., Adams, D.B., Barnett, J.P., Hinch, G.N., Munro, R.K., & Davies, H.I. (1992). The link between immune responses and behavior in sheep. In: A.L. Husband, (Ed.) Behaviour and immunity (pp. 23-41). Boca Raton, Florida: CRC Press.
Huitema, B.E. (1980). The analysis of covariance and alternatives. New York: J. Wiley & Sons.
Ludány, A., Gallyas, F., Gaszner, B., Andrásfalvy, B., Szücs, G., & Kellermayer, M. (1993). Skimmed-milk blocking improves silver post-intensification of peroxidase-diaminobenzidine staining on nitrocellulose membrane in immunoblottig. Electrophoresis, 14, 78-80.
Maier, S.F., & Watkins, L.R. (1998). Cytokines for psychologists: implications of bi-directional immune-to-brain communication for understanding behavior, mood, and cognition. Psychological Reviews, 105, 83-107.
Simmel,E.C. & Bagwell, M. (1983). Genetics of exploratory behavior and activity. In: J.L. Fuller & E.C. Simmel, (Eds.) Behavior genetics: Principles and applications (pp. 89-115). Hillsdale, NJ: Erlbaum.
Vidal, J. (1999). Influence of the specific immune response on some consistent murine behaviors. Journal of General Psychology, 126, 205-216.
von Horsten, S., Exton, M.S., Voge, J., Schult, M., Nagel, E., Schmidt, R.E., Westermann, J., & Schedlowski, M. (1998). Cyclosporine A affects open field behavior in DA rats. Pharmacology, Biochemistry and Behavior, 60, 71-76.
Wilcox, R.R. (1987). New statistical procedures for the social sciences. Modern solutions to basic problems. Hillsdale, NJ: Erlbaum.
Winkelstein, A. (1994). Immunosuppressive therapy. In D.P. Stites, A.I. Terr, & T.G. Parslow (Eds.), Basic and clinical immunology (8th ed.) (pp.765-780). London: Prentice Hall.
Zacharko, R.M., Zalcman, S., MacNeil, G., Andrews, M., Mendella, P.D., & Anisman, H. (1997). Differential effects of immunologic challenge on self-stimulation from the nucleus accumbens and the substantia nigra. Pharmacology, Biochemistry, and Behavior, 58, 881-886.
Table 1
Sequence of operations in female mice
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Table 2
Sequence of operations in male mice
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
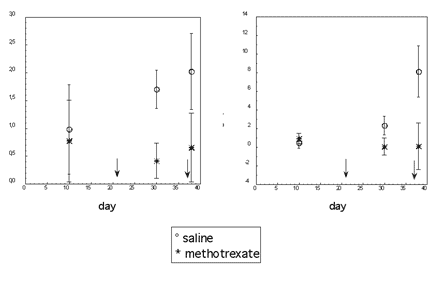
Figure 1. Antibody response in female mice injected with methotrexate (N = 12) or with saline (N = 10). Arrows indicate beginning and end of methotrexate administration; the drug was injected daily at 3 mg/kg. Bars are 95% confidence intervals. Antibodies on day 10 are antibodies to goat albumin (2mg/mouse) injected on day 2. Antibodies on day 30 are antibodies to rat erythrocytes (4.8 × 107 /mouse) injected on day 21, and antibodies on day 38 are antibodies to a second injection of the same antigen (4.2 × 107 erythrocytes/mouse) on day 30.
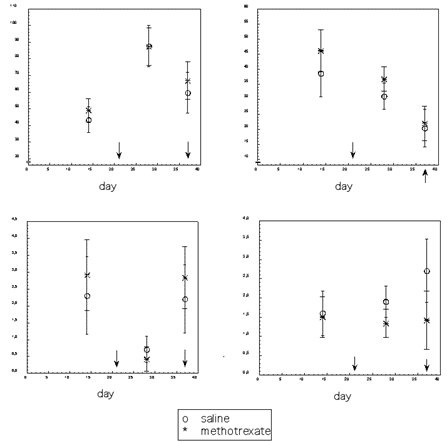
Figure 2. Behaviors in the open field of female mice injected with methotrexate (N = 12) or with saline (N = 10). Arrows indicate beginning and end of methotrexate administration; the drug was injected daily at 3 mg/kg. Bars are 95% confidence intervals.
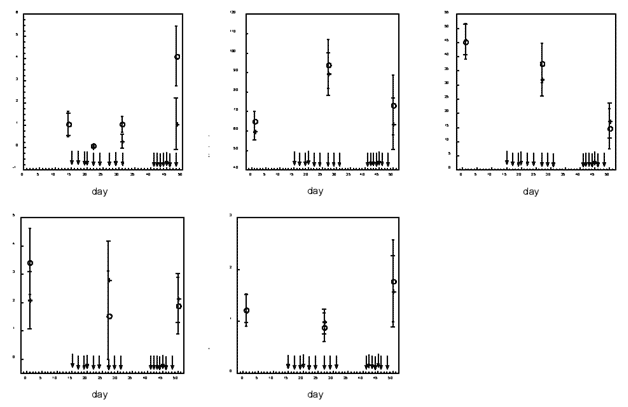
Figure 3. Antibody response and behaviors of male mice injected with saline (N = 9) or with methotrexate (N = 12). Each arrow indicates a methotrexate injection (3 mg/kg/injection). Bars are 95% confidence intervals. Antibodies on day 14 are antibodies to goat albumin (2mg/mouse) injected on day 6. Antibodies on days 22 and 31 are antibodies to rat erythrocytes (3.9 × 107 /mouse) injected on day 15, whereas antibodies on day 48 are antibodies to a second injection of the same antigen (4.4 × 107 erythrocytes/mouse) on day 41.
o: saline; *: methotrexate
Jose Vidal
Departamento de Personalidad,
Facultad de Psicologia,
Universidad de Barcelona;
Passeig de la Vall d'Hebron,
171; 08035 Barcelona, Spain
jvidal@psi.ub.es
Accepted: November 13, 2002
Revised: November 20, 2002